






乙烯是植物最经典的气体激素之一,参与生长发育与逆境响应;在拟南芥中,乙烯由内质网膜上的5个乙烯受体感知,并按序列特征分成I亚家族(ETR1、ERS1)与II亚家族(ETR2、ERS2、EIN4)。另一方面,Ca²⁺是植物细胞里最重要的第二信使之一,很多刺激(包括激素)都会引发胞质Ca²⁺瞬变;早在30多年前就有人发现乙烯响应需要Ca²⁺,也有人通过膜片钳观察到乙烯能激活Ca²⁺通道并抬升胞质Ca²⁺,但是具体乙烯诱导Ca²⁺信号的分子来源长期未知。

乙烯直接影响胞质Ca²⁺浓度,而且ETR1/ERS1缺失会让这波信号“变弱”
首先作者用表达aequorin的拟南芥幼苗检测胞质Ca²⁺:乙烯(ethephon)处理能显著提高钙离子浓度;加入内源Ca²⁺通道阻断剂Ruthenium Red后,这个提升被压到对照的约45%,提示乙烯诱导的Ca²⁺上升至少部分来自内源Ca²⁺通道。更关键的是:对比多种受体突变体,乙烯诱导的[Ca²⁺]cyt上升在etr1和etr1 ers1双突变体中降低,而在一些II亚家族相关突变体中基本正常,提示乙烯诱导的Ca²⁺信号与I亚家族受体ETR1/ERS1密切相关。
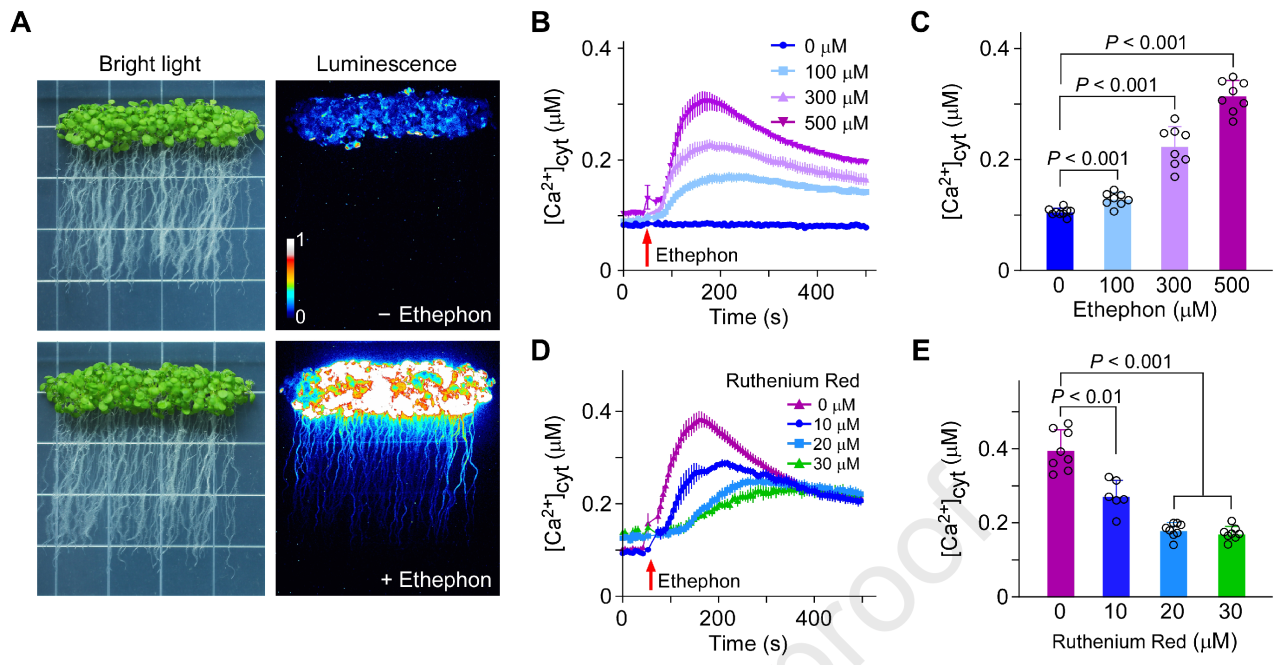
图1. 乙烯提高胞质Ca²⁺浓度, ETR1/ERS1缺失降低相应
电生理“坐实”:乙烯受体I亚家族ETR1/ERS1能导Ca²⁺,II亚家族不导;La³⁺能阻断
随后作者把5个拟南芥乙烯受体分别在爪蟾卵母细胞表达并做膜片钳实验:ETR1与ERS1出现内向Ca²⁺电流;加入外源Ca²⁺通道阻断剂La³⁺后电流被消除,具有典型的钙离子通道特征;而ETR2/ERS2/EIN4检测不到相应Ca²⁺电流——也就是“I亚家族能导钙,II亚家族不导”。
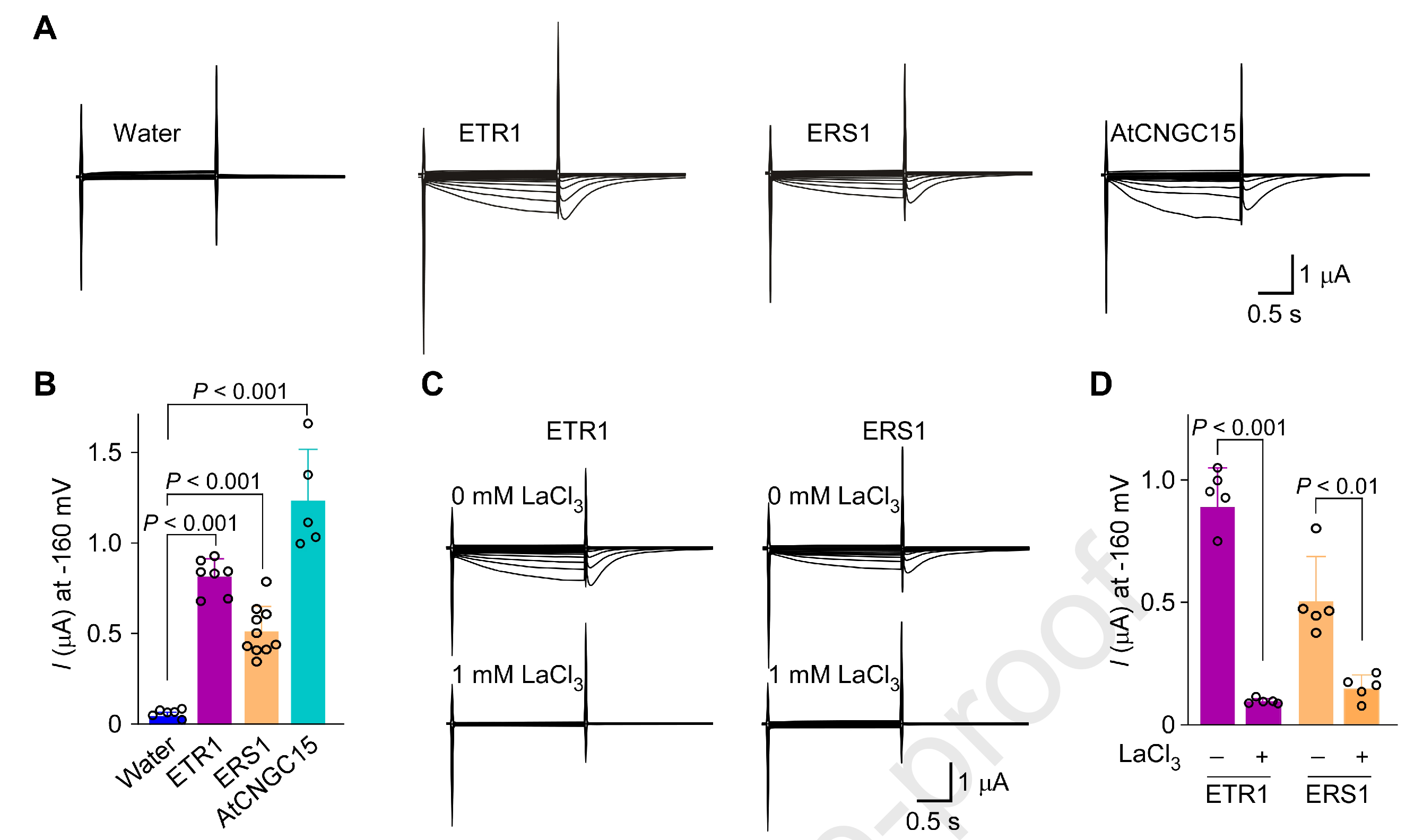
图2. 乙烯受体I亚家族ETR1/ERS1有钙离子通道特征
“结构门槛”:受体二聚化是导钙的必要条件
ETR1以同源二聚体作为基本功能单元。作者把ETR1 N端参与二聚化的Cys4/Cys6突变为Ser以破坏二聚化:结果二聚体只在WT中形成,而突变体既不形成二聚体,也丧失Ca²⁺通透性。
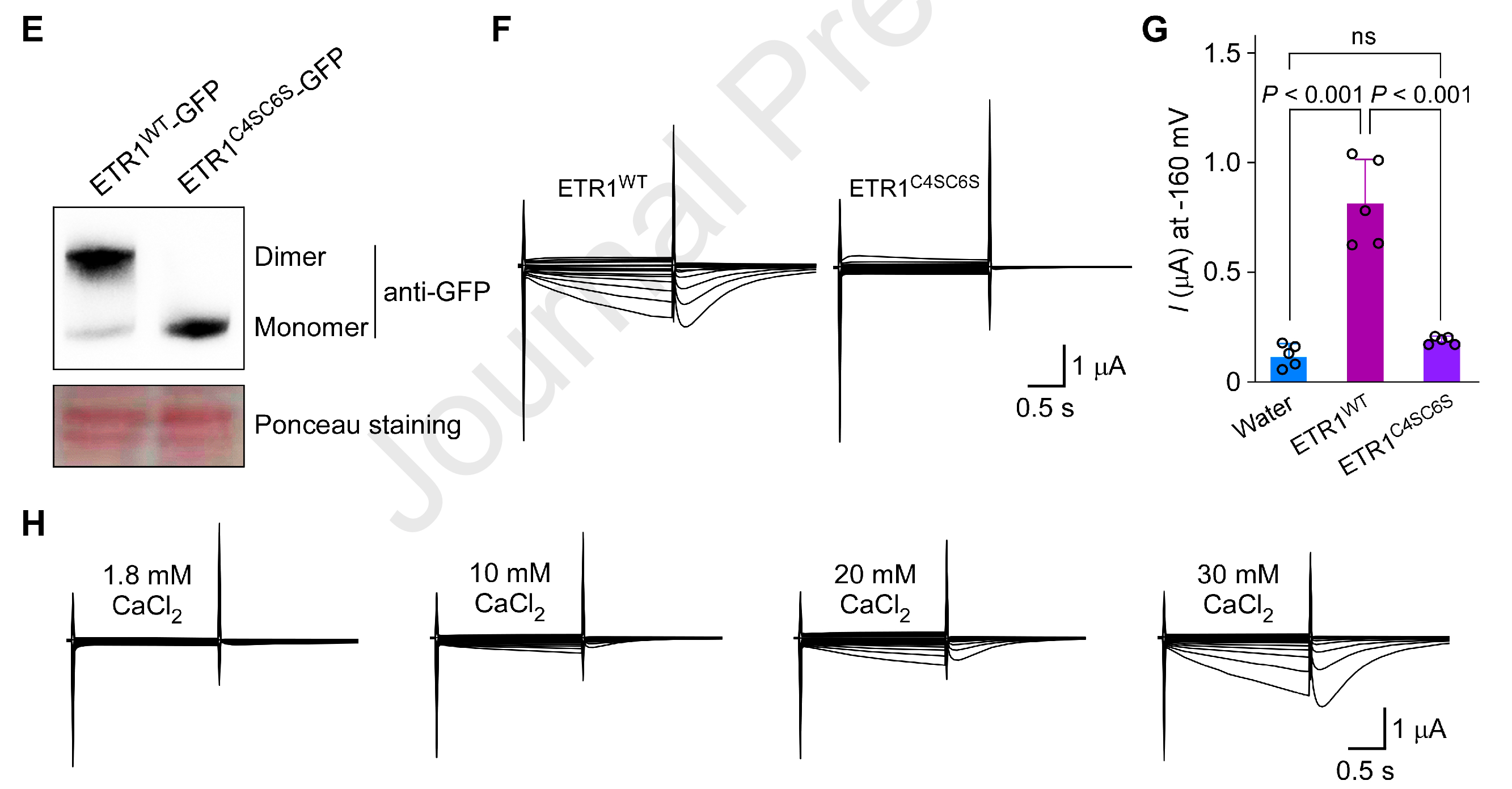
图3. 乙烯受体钙通道活性需要二聚体形式
不止拟南芥:ETR1的导钙能力在“绿色谱系”里高度保守
作者挑选了从藻类到苔藓、地钱、卷柏,再到单子叶/双子叶等多个类群的ETR1同源蛋白,在卵母细胞中测试后发现:所有被检测的ETR1同源物都能产生清晰的Ca²⁺电流,提示这种功能可能起源很早,并在绿色谱系中保留下来。
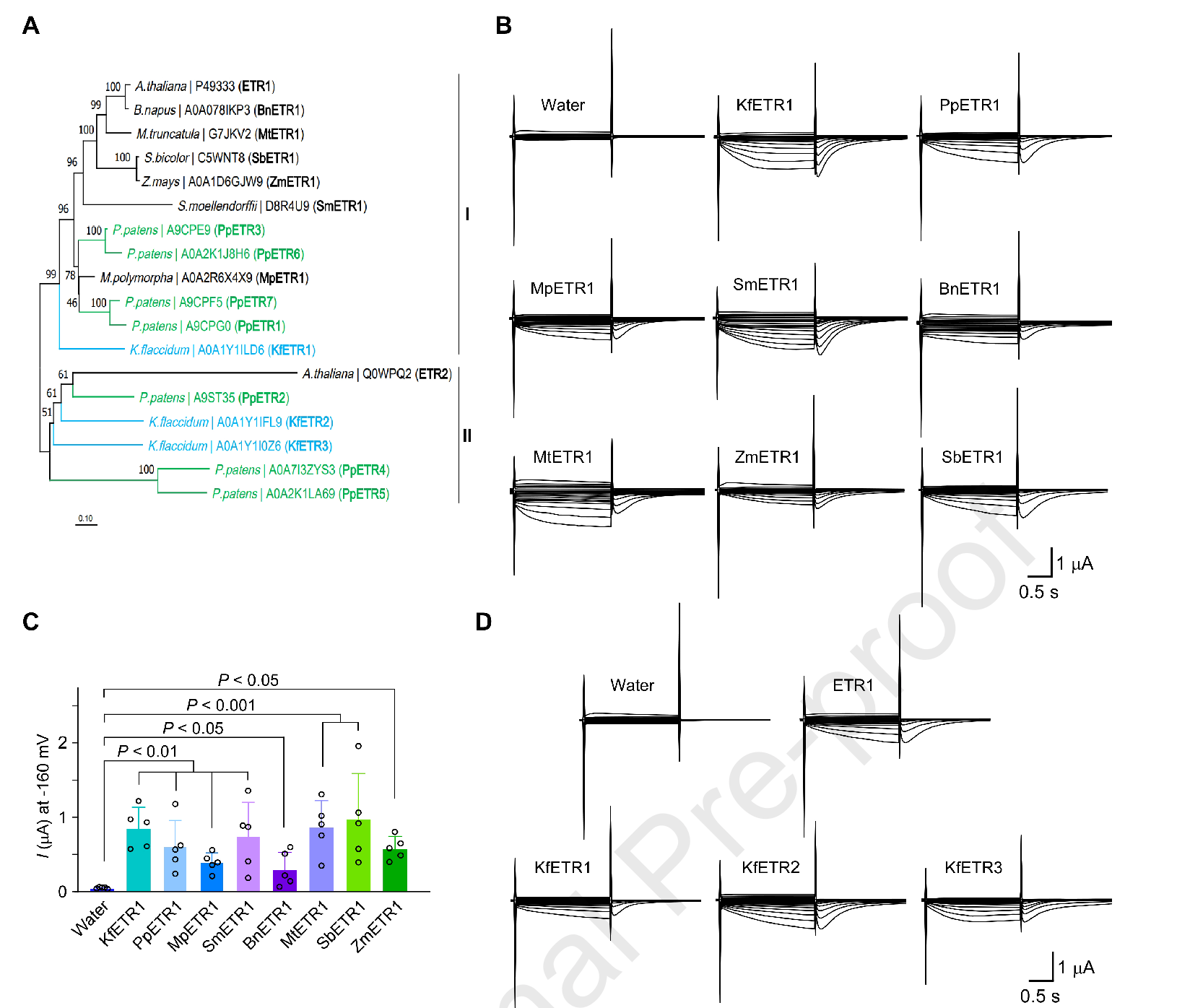
图4. ETR1的导钙能力在“绿色谱系”里高度保守
乙烯配体结合可增强受体介导的 Ca²⁺通透性/电导
一个很有意思的点:作者发现ethephon能进一步增强ETR1在卵母细胞中的Ca²⁺电流;但当乙烯结合位点突变(Cys65Ser)后,ethephon对电流的增强效应消失,提示这是配体依赖的调控。同样的现象也出现在藻类的同源受体上,说明“乙烯调控导钙”可能在登陆之前就已存在。
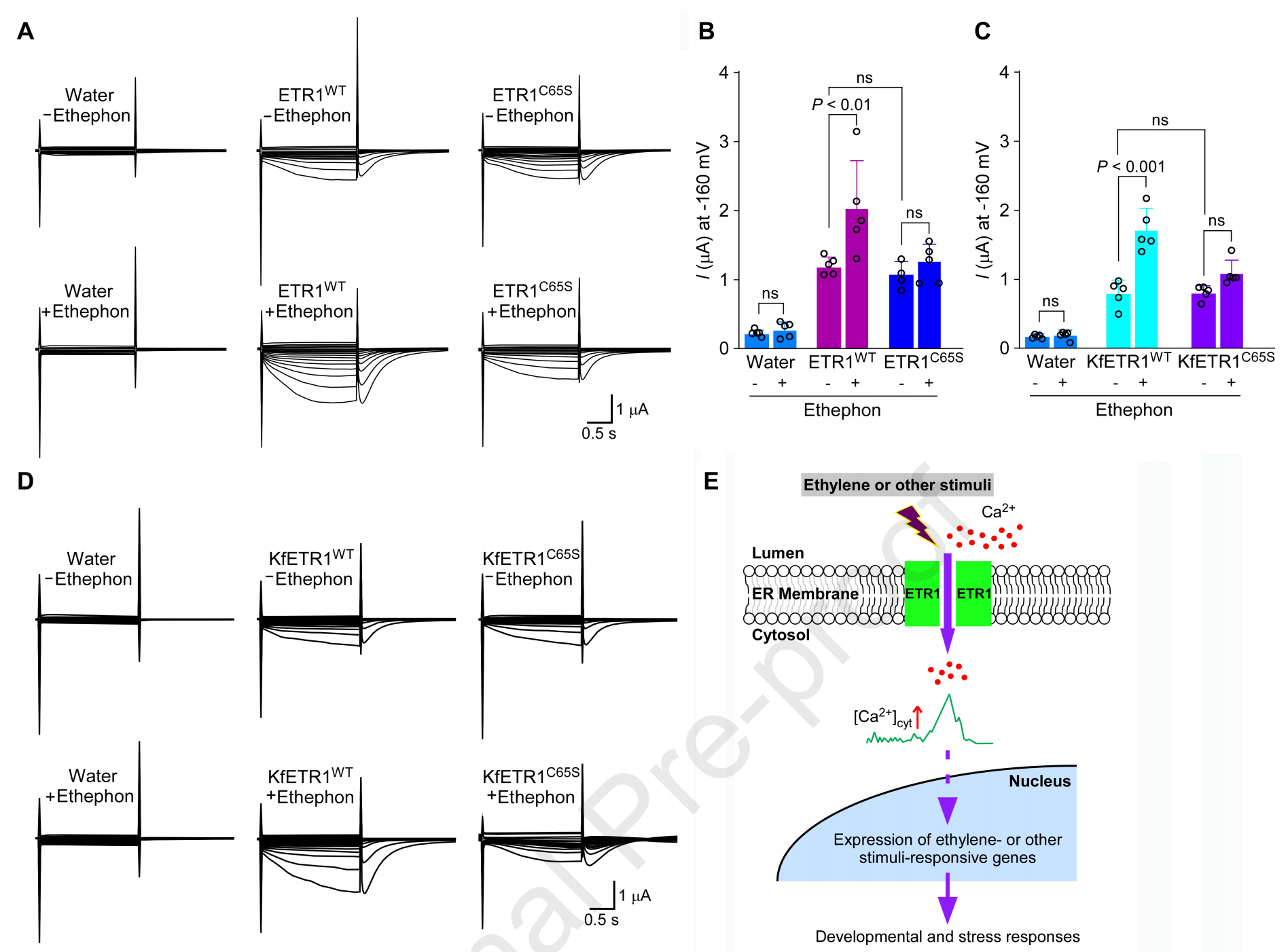
总之,本研究揭示I亚家族乙烯受体ETR1/ERS1具有Ca²⁺通透性,乙烯可增强其介导的Ca²⁺内向电流;缺失ETR1/ERS1会削弱乙烯诱导的胞质Ca²⁺升高。多种藻类与陆生植物ETR1同源蛋白同样导钙,提示该功能在绿色谱系中保守。该工作将乙烯受体从“信号感受器”拓展为潜在的Ca²⁺通道组分,建立乙烯—Ca²⁺耦联的分子基础,为解析激素信号编码与胁迫响应提供新机制与干预靶点。

