






自然进化通常难以直接研究,因为历史事件往往只发生一次且受随机突变和偶然性影响,而趋同进化为探讨进化可重复性提供了重要线索。蛋白质趋同已在残基、功能和结构域层面被研究,但整体三维结构的重复性趋同仍知之甚少。基因融合可改变蛋白质结构域组合,促进全局结构演化,并可多次独立发生。醛脱氢酶/醇脱氢酶(ALDH/ADH)融合酶是研究这一现象的理想模型,例如已知 AdhE 家族通过基因融合形成螺旋同源聚合物,促进底物通道化和双步反应效率。

BdhE 是一种 双功能脱氢酶家族(Bifunctional dehydrogenase E),由 醛脱氢酶(ALDH)和醇脱氢酶(ADH)基因独立融合形成,但进化上与已知的AdhE 家族完全不同。通过在 45,555 个细菌基因组中搜索 AdhE 序列,研究者发现了一组与 AdhE 序列相似度低(<30%)<>但覆盖度高(>90%)的基因,这些基因编码的蛋白结构类似AdhE,却在系统发育树上形成独立的单系群,支持 BdhE 与 AdhE 来源于不同的融合事件(图1c)。BdhE 的 ALDH 和 ADH 域分别与各自的近亲基因簇相关,但与AdhE 的对应域不在同一单系群(图1d)。由于功能和结构上与 AdhE 类似,BdhE 成为研究趋同基因融合及结构演化的理想模型。
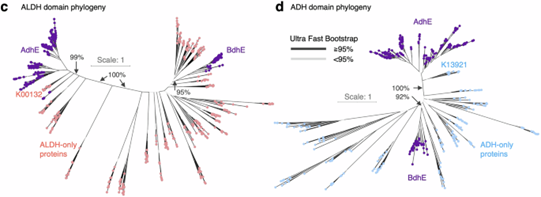
图 1. AdhE 与 BdhE 具有相似但进化独立的结构域架构
BdhE 与 AdhE 虽然进化独立,但两者的 ALDH 和 ADH 域显示了相似的酶活性,包括乙醇氧化和乙酰辅酶 A 还原,以及胆碱氧化(图2a、b)。实验表明,BdhE 的反应速率低于 AdhE,但功能方向一致。宏观进化分析显示,两者在细菌中的系统发育分布互不重叠,表明其功能冗余且不适合共存(图2c)。环境分析则进一步显示,BdhE 富集于水-土混合和盐水环境,而 AdhE 富集于人类肠道及相关环境,两者可能在不同生态位中承担类似的厌氧发酵功能,同时BdhE 可能参与高渗环境下的甜菜碱合成。
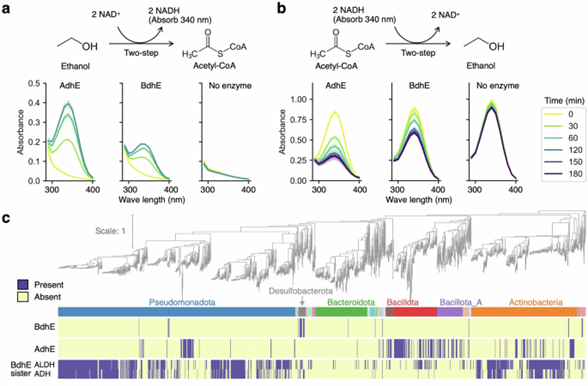
图 2. AdhE 与 BdhE 具有相似酶活性,但系统发育及生态分布不重叠
BdhE 和 AdhE 虽然酶活性相似,但多聚体结构不同:BdhE 形成稳定的环状四聚体,而 AdhE 形成螺旋状聚合物(图 3a)。冷冻电镜显示,两者的多聚体均由弯曲二聚体结构单元组成,但二聚体的弯曲角度不同导致整体结构差异(图 3a, c, d)。BdhE 的环状四聚体比 AdhE 的螺旋体更紧凑,无法形成开放的螺旋结构(图 3b)。AlphaFold 能较准确预测 BdhE 的二聚体和四聚体,但若无实验验证,环状结构可能会被误判为 AdhE。这一发现表明,独立融合的基因可通过类似二聚体单元演化出不同的三维多聚体构象。
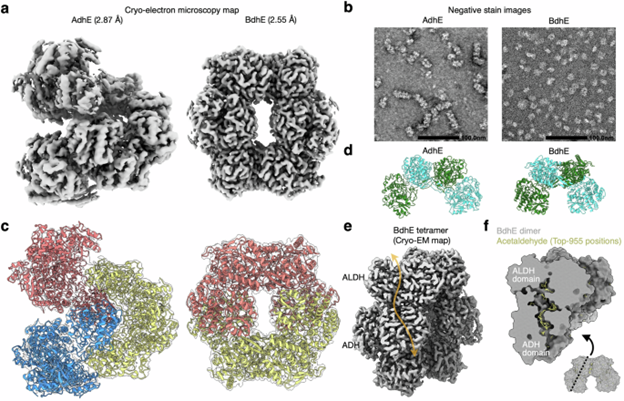
图 3. AdhE 与 BdhE 的冷冻电镜结构揭示结构域间相互作用的趋同性
BdhE 和 AdhE 虽然独立进化,但都形成了二聚体结构,实现 ALDH 与 ADH 域之间的底物通道化。冷冻电镜和分子对接显示,两者的二聚体都能稳定传递底物,但通道结构存在差异(图 3e, f)。界面分析发现,AdhE 和 BdhE 的 ALDH–ADH 接触由不同的非同源环状结构(loops)形成,这些环在各自的近缘 ALDH 或 ADH 基因中不存在,表明通道界面是通过独立的环延长趋同演化而来 (图4)。BdhE 二聚体的通道存在间隙,但在四聚体中由另一二聚体部分填充,使其在防止底物泄漏方面更有效。总体来看,AdhE 和 BdhE 在结构层面趋同进化了底物通道界面,但机制各异。
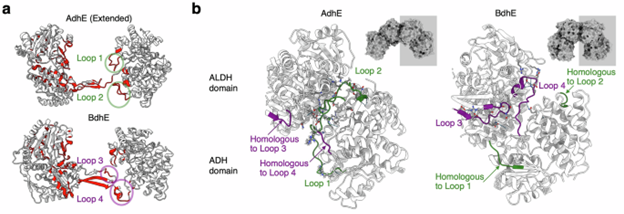
图 4. 趋同进化的 AdhE 与 BdhE 多聚体具有不同特性
之后,研究人员推测AdhE 和 BdhE 的独立基因融合可能受其祖先 ALDH 和 ADH 基因在基因组中邻近位置的促进,这使得单域基因通过小片段缺失容易融合。邻近基因对通常功能相关并共转录,但在融合前并未形成跨域相互作用。融合后的 BdhE 和 AdhE 通过各自特异的环结构趋同获得 ALDH–ADH 接触,实现底物通道化。祖先单域基因的同源二聚体相互作用为多聚体形成提供基础,而跨域相互作用则是融合后独立演化获得的。
最后,研究者还提出了 AdhE/BdhE 的进化模型(图5):最初,编码 ALDH 和 ADH 的基因分别形成同源二聚体或四聚体,部分基因随后在基因组中相邻并发生融合。融合后的蛋白最初没有跨域相互作用,随后通过环延长演化出 ALDH–ADH 接触界面,最终驱动 AdhE 形成可变螺距的螺旋聚合物,BdhE 形成环状四聚体,实现二聚体界面趋同而整体多聚体结构分化。
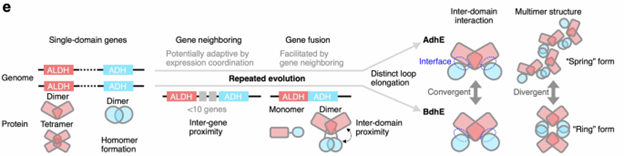
图5. AdhE 与 BdhE 的趋同进化模型

