






植物为了生长发育,需要面对来自外界的各种挑战,免疫系统便是它们的“防线”。在植物的免疫系统中,CERK6是非常关键的受体激酶,它识别并响应几丁质(几乎所有真菌的细胞壁成分),启动植物的免疫反应,帮助植物抵御病菌入侵。但植物不仅仅要防守,还要与根瘤菌这种“友好”的细菌建立共生关系。根瘤菌通过与豆科植物的根部合作,帮助植物固定空气中的氮,从而提高土的肥力。这个过程中,结瘤因子受体NFR1担当了重要角色,它识别根瘤菌发出的“邀请信号”(诺多因子,Nod因子),启动共生反应,帮助植物建立“友谊”。尽管CERK6和NFR1在功能上差异巨大,但它们的结构却非常相似。那么,是什么使得它们能在面临相似的信号时,做出完全不同的反应呢?
2025年11月5日,丹麦奥胡斯大学的Kasper Røjkjær Andersen和Simona Radutoiu团队在顶级期刊《Nature》上发表了一篇题为“Two residues reprogram immunity receptors for nitrogen-fixing symbiosis”的研究。该研究首次揭示了植物免疫受体NFR1内的两个关键氨基酸残基如何控制受体在免疫反应和固氮共生之间的功能切换。通过对NFR1与CERK6受体的结构和功能比较,研究者发现,两个残基不仅决定了植物如何响应病原挑战,还能够通过小的结构改变,重编程这些受体以支持根瘤菌的固氮共生。这一发现为理解植物免疫与共生之间的分子机制提供了新的视角,并为非豆科作物的固氮能力工程改良提供了潜在的突破口。

“共生决定因子1”(SD1)区域在共生信号
中的核心作用
研究者首先构建了多个NFR1和CERK6的嵌合受体变体,以测试这些受体在免疫反应与共生反应中的功能差异(图1)。具体来说,这些变体结合了NFR1的外细胞结构域CERK6的激酶结构域,帮助定位哪些区域对共生信号的启动至关重要。这些嵌合受体变体被表达在百脉根(Lotus japonicus)的nfr1突变体中。功能测试表明,NFR1受体的嵌合变体和含有Symbiosis Determinant 1(SD1)区域的CERK6变体能够成功启动结节的形成,即固氮结节。而没有SD1的CERK6受体变体,则无法启动结节形成,无法激活固氮反应。SD1不仅是启动共生反应的必要区域,它还决定了NFR1和CERK6在免疫与共生信号中的功能分化,明确SD1在调控共生信号启动中的核心地位。
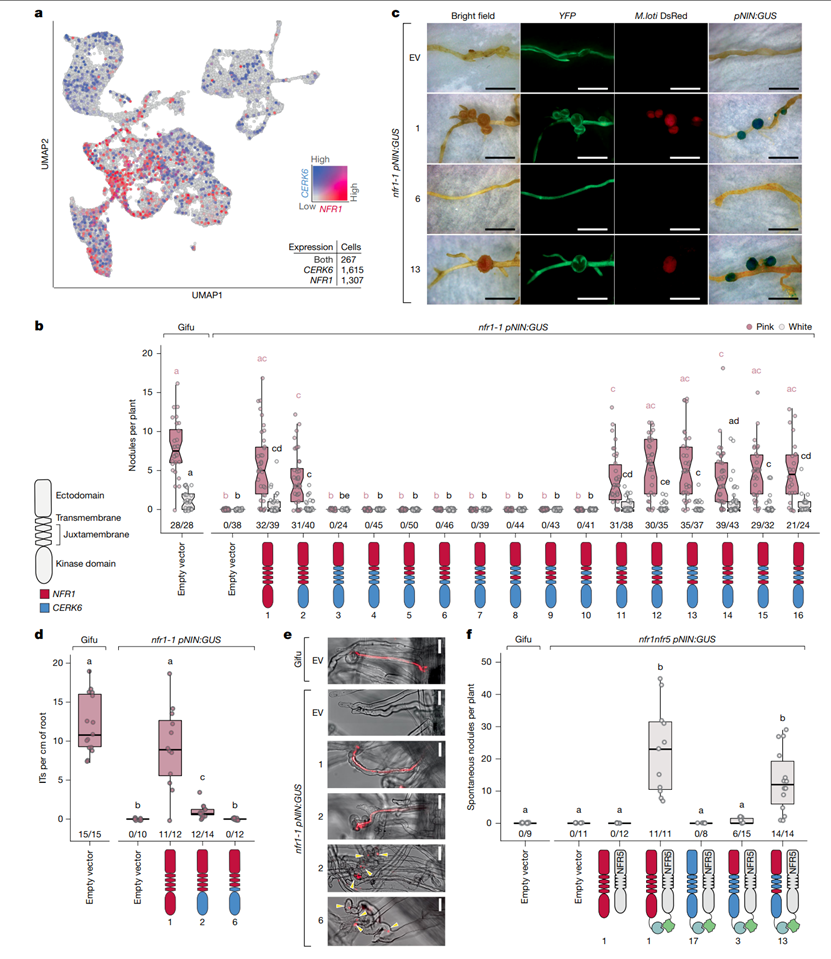
图1. Symbiosis Determinant 1(SD1)控制根瘤共生
结构解析揭示SD1的空间布局及其对信号
传导的影响
随后研究人员解析了CERK6和LYK3的激酶结构,旨在从结构层面理解SD1的作用(图2)。通过对比两者的结构,研究人员试图揭示SD1在分子中的定位及其如何通过与其他分子相互作用影响信号传导。结果显示,SD1位于激酶结构域的跨膜邻区(灰色部分),并且该区域的氨基酸残基暴露在受体的表面。这种表面暴露的结构特性使SD1能够与其他分子(如NFR5)进行相互作用,从而调节共生信号的传递。
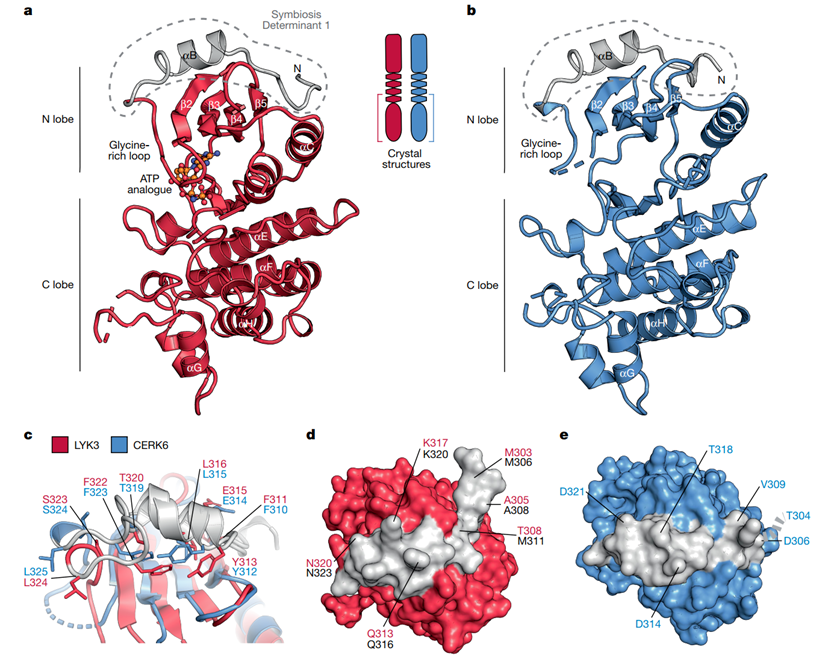
图2. LYK3与CERK6激酶结构域的晶体结构
激酶表面区域的差异:免疫与共生信号
的分离机制
为了进一步探讨免疫反应与共生反应之间的分子机制,研究人员通过交换CERK6和NFR1受体激酶的表面区域,测试这些变体在两种反应中的表现。这个实验旨在揭示受体激酶的哪些区域控制着免疫与共生信号的启动。研究发现,NFR1和CERK6的C末端区域以及激活环区域对于免疫和共生信号的分离至关重要。当CERK6和NFR1的这些区域发生交换时,受体的功能发生变化,表明这些表面区域在调节信号通路的特异性方面起着决定性作用。结果表明,CERK6受体通过其C末端区域启动免疫反应,产生活性氧(ROS)。而NFR1则无法激活免疫反应,进一步确认了免疫信号和共生信号在分子水平上的独立性。
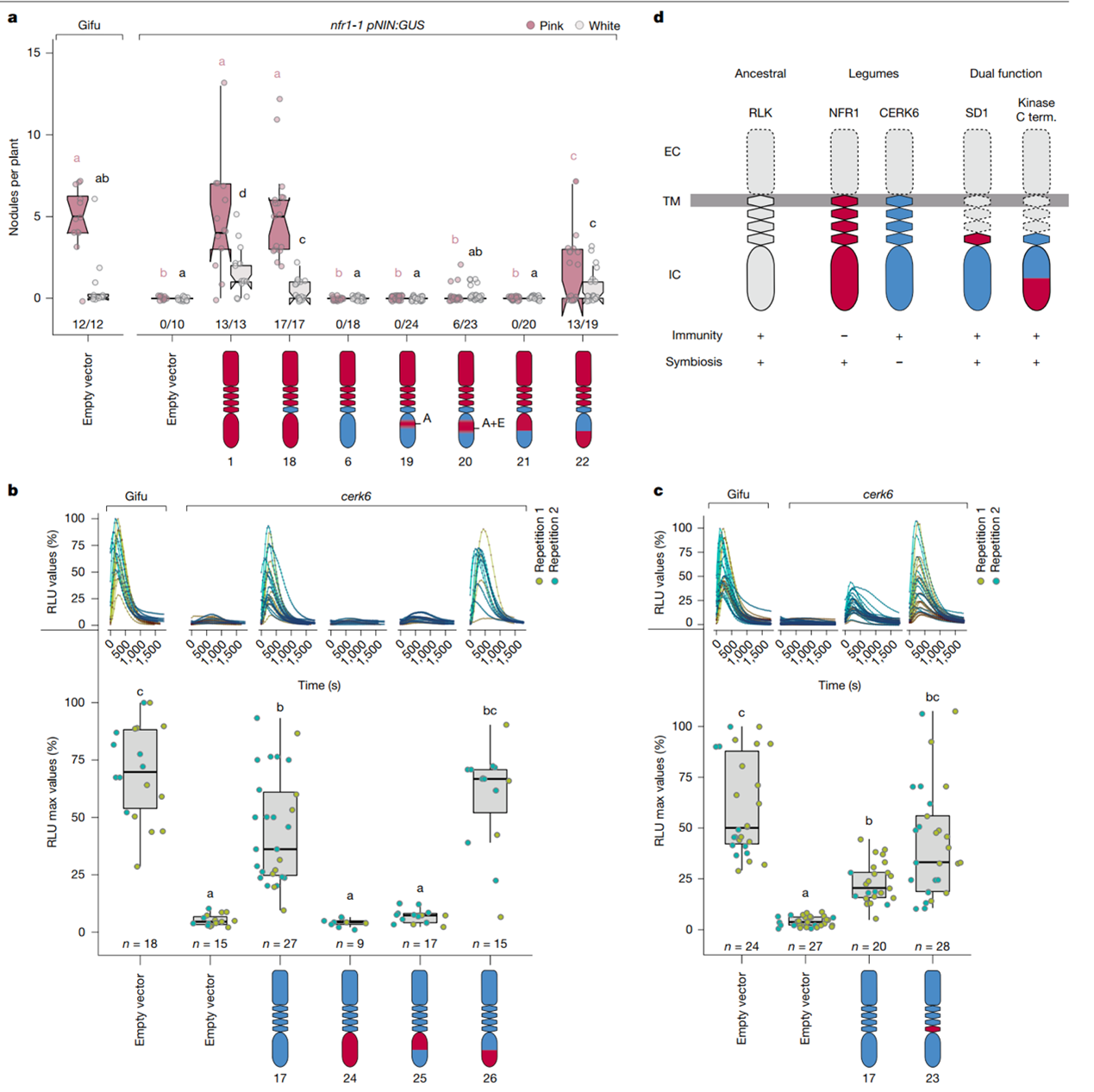
图3. NFR1和CERK6激酶结构域的差异
SD1使CERK6和RLK4受体具备共生信号
之后研究人员进一步探索了SD1区域在植物受体中的功能作用,展示了SD1如何通过结构改变,使得CERK6和RLK4受体从激活免疫反应转变为启动共生信号。通过对NFR1、CERK6和RLK4的SD1区域氨基酸序列进行比对,发现SD1区域在NFR1和CERK6中的氨基酸残基存在显著差异,这些差异与受体的功能密切相关。研究通过引入SD1,成功使CERK6和RLK4受体从激活免疫信号(如ROS生成)转变为启动共生信号,从而诱导根瘤共生反应并形成结节。特别地,T304M和D306A这两个氨基酸残基的突变使CERK6能够在百脉根(Lotus japonicus)中成功启动结节的形成,证明了这两个氨基酸在共生反应中的关键作用。SD1不仅能转换免疫信号为共生信号,还能抑制免疫反应,确保免疫与共生信号之间的平衡。这表明,SD1在调节免疫和共生反应的功能转换中起着核心作用。值得注意的是,SD1不仅在豆科植物中发挥作用,还能在非豆科植物的受体(如RLK4)中表现出类似的功能,进一步为非豆科作物的固氮共生提供了潜在的工程化改良路径。
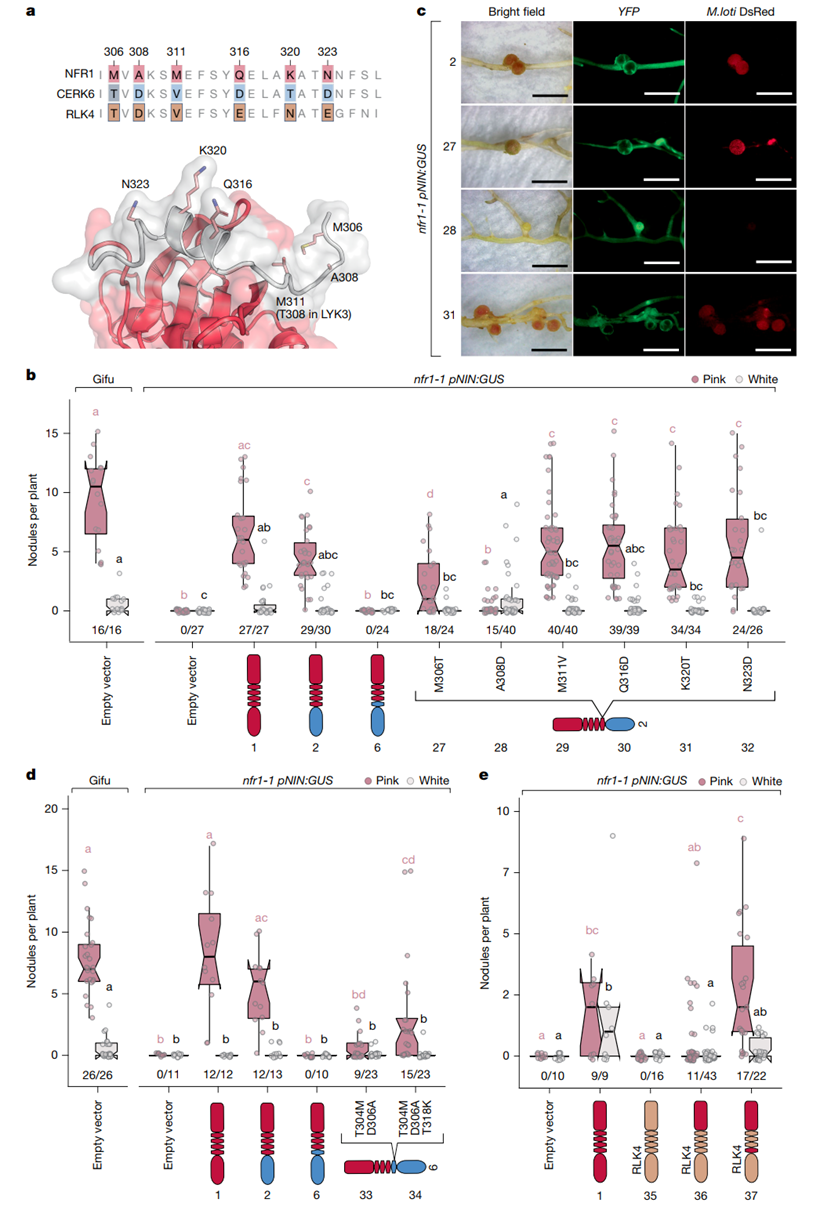
图4. SD1区域在调控受体功能切换中的关键作用
总之,本文揭示了Symbiosis Determinant 1(SD1)区域在植物免疫受体功能切换中的关键作用。研究发现,SD1通过两个关键氨基酸(T304M和D306A)突变,能将CERK6和RLK4等受体从激活免疫反应转变为启动根瘤共生信号,成功诱导结节形成。SD1不仅能在豆科植物中发挥作用,还能在非豆科植物的受体中表现出类似功能,提供了非豆科作物固氮共生的潜力。此外,SD1调控免疫与共生信号之间的平衡,避免免疫反应过度激活。研究为植物受体的功能工程化提供了新的思路,并为农业中固氮共生的应用开辟了新的路径。

